Строение коллагена – особенности аминокислотного состава, первичной и пространственной структуры. Особенности биосинтеза и созревания коллагена. Роль аскорбиновой кислоты в созревании коллагена.
Коллаген — Википедия
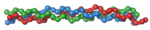
Коллаге́н — фибриллярный белок, составляющий основу соединительной ткани организма (сухожилие, кость, хрящ, дерма и т. п.) и обеспечивающий её прочность и эластичность. Коллаген обнаружен у животных; отсутствует у растений, бактерий, вирусов, простейших и грибов[1]. Коллаген — основной компонент соединительной ткани и самый распространённый белок у млекопитающих[2], составляющий от 25 % до 45% белков во всём теле. Синтез коллагена очень энергозатратный и происходит только у животных, которые используют свободный кислород. Появление коллагена позволило создать скелет, как внешний и внутренний, и резко увеличить размеры животных во время Кембрийского взрыва.
Учёные десятилетиями не могли понять молекулярное строение коллагена. Первое доказательство того, что коллаген имеет постоянное строение на молекулярном уровне, было представлено в середине 1930-х годов. С того времени много выдающихся учёных, включая Нобелевских лауреатов, таких как Фрэнсис Крик, Лайнус Полинг, Александр Рич, Ада Йонат, Хелен Берман, Вилеайнур Рамачандран, работали над строением мономера коллагена.
Несколько противоречащих друг другу моделей (несмотря на известное строение каждой отдельной пептидной цепи) дали дорогу для создания троично-спиральной модели, объяснившей четвертичное строение молекулы коллагена.
Продуктом денатурации коллагена является желатин. Температура денатурации макромолекулы коллагена близка к температуре фибриллогенеза. Это свойство молекулы коллагена делает её максимально чувствительной к мутационным заменам.
Фибриллогенез — образование коллагеновых волокон в соединительной ткани путём сборки или объединения в пучки фибрилл — тонких белковых нитевидных структур внутри клетки и тканей человеческого организма. Фибриллогенез имеет важное значение в процессе приживления имплантанта и создания на его основе прочной, правильно функционирующей жевательной системы. Чем прочнее созданные в процессе фибриллогенеза коллагеновые волокна, тем прочнее соединительная ткань.
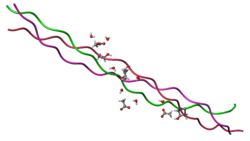
Молекула коллагена представляет собой левозакрученную спираль из трёх α-цепей. Такое образование известно под названием
Для первичной структуры белка характерно высокое содержание глицина, низкое содержание серосодержащих аминокислот и отсутствие триптофана. Коллаген относится к тем немногим белкам животного происхождения, которые содержат остатки нестандартных аминокислот: около 21 % от общего числа остатков приходится на 3-гидроксипролин, 4-гидроксипролин и 5-гидроксилизин[4]. Каждая из α-цепей состоит из триад аминокислот. В триадах третья аминокислота всегда глицин, вторая — пролин или лизин, первая — любая другая аминокислота, кроме трёх перечисленных
Коллаген существует в нескольких формах. Основа строения всех видов коллагена является схожей. Коллагеновые волокна образуются путём агрегации микрофибрилл, имеют розовый цвет при окраске гематоксилином и эозином и голубой или зелёный при различных трёххромных окрасках, при импрегнации серебром окрашиваются в буро-жёлтый цвет.
Тропоколлагены (структурные единицы коллагена) спонтанно объединяются, прикрепляясь друг к другу смещёнными на определённое расстояние концами, образуя в межклеточном веществе более крупные структуры. В фибриллярных коллагенах молекулы смещены относительно друг друга примерно на 67 нм (единица, которая обозначается буквой «D» и меняется в зависимости от состояния гидратации вещества). В целом каждый D-период содержит четыре целых и часть пятой молекулы коллагена. Величина 300 нм, поделённая на 67 нм (300:67), не даёт целого числа, и длина молекулы коллагена разделена на непостоянные по величине отрезки D. Следовательно, в разрезе каждого повтора D-периода микрофибриллы есть часть, состоящая из пяти молекул, называемая «перекрытие», и часть, состоящая из четырёх молекул — «разрыв». Тропоколлагены к тому же скомпонованы в шестиугольную или псевдошестиугольную (в поперечном разрезе) конструкцию, в каждой области «перекрытия» и «разрыва».
Внутри тропоколлагенов существует ковалентная связь между цепями, а также некоторое непостоянное количество данных связей между самими тропоколагеновыми спиралями, образующими хорошо организованные структуры (например, фибриллы). Более толстые пучки фибрилл формируются с помощью белков нескольких других классов, включая другие типы коллагенов, гликопротеины, протеогликаны, использующихся для формирования различных типов тканей из разных комбинаций одних и тех же основных белков. Нерастворимость коллагена была препятствием к изучению мономера коллагена, до того момента как было обнаружено, что возможно извлечь тропоколлаген молодого животного, поскольку он ещё не образовал сильных связей с другими субъединицами фибриллы. Тем не менее, усовершенствование микроскопов и рентгеновских аппаратов облегчили исследования, появлялось всё больше подробных изображений структуры молекулы коллагена. Эти поздние открытия очень важны для лучшего понимания того, как структура коллагена влияет на связи между клетками и межклеточным веществом, как ткани меняются во время роста и регенерации, как они меняются во время эмбрионального развития и при патологии.
Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена. Коллагеновые волокна — это пучки фибрилл.
Пищевая промышленность[править | править код]
С точки зрения питания, коллаген и желатин являются белками низкого качества, так как они не содержат всех незаменимых аминокислот, необходимых человеку — это неполноценные белки. Производители основанных на коллагене пищевых добавок утверждают, что их продукты могут улучшить качество кожи и ногтей, а также здоровье суставов
Относительно дешёвые, часто предлагаемые сегодня на рынке под видом источника свободных аминокислот гидролизаты коллагена не всегда способны удовлетворить потребности человека в свободных аминокислотах, так как эти продукты не содержат готовые к усвоению аминокислоты, а являются лишь частично «переваренными» экстрактами суставных тканей млекопитающих, птиц или обитателей моря. Например, гидролизаты коллагена почти полностью лишены аминокислоты L-глютамина, не отличающейся стойкостью к термическому воздействию и долгому хранению сырья, большая часть глютамина разрушается уже на первых этапах хранения и переработки сырья, имеющийся небольшой остаток практически полностью распадается во время термической экстракции хрящевой ткани.
Наиболее качественными источниками аминокислот являются препараты, содержащие так называемые «свободные аминокислоты». Так как именно свободные аминокислоты являются практически готовыми к усвоению, организму не нужно тратить пищеварительные ферменты, время и энергию на их переваривание. Они способны в кратчайшие сроки поступить в кровь и, будучи доставленными ею к местам, нуждающимся в дополнительном синтезе коллагена, тут же включаются в его формирование
Косметические средства[править | править код]
Коллаген входит в состав косметических средств для:
- Образования воздухопроницаемого, влагоудерживающего слоя на поверхности кожи, обладающего пластифицирующими (разглаживающими) свойствами, со свойствами влажного компресса;
- Продления действия экстрактов, масел и др. в составе косметических композиций;
- Придания блеска волосам, создания коллагенового (защитного) слоя на поверхности волос.
Научные исследования[править | править код]
В 2005 году учёным удалось выделить коллаген из сохранившихся мягких тканей тираннозавра[5][нет в источнике] и использовать его химический состав как ещё одно доказательство родства динозавров с современными птицами
Научные исследования в медицине[править | править код]
Синтез коллагена — сложный ферментативный многостадийный процесс, который должен быть обеспечен достаточным количеством витаминов и минеральных элементов. Синтез протекает в фибробласте и ряд стадий вне фибробласта. Важный момент в синтезе — реакции гидроксилирования, которые открывают путь дальнейшим модификациям, необходимым для созревания коллагена. Катализируют реакции гидроксилирования специфические ферменты. Так, образование 4-оксипролина катализирует пролингидроксилаза, в активном центре которой находится железо. Фермент активен в том случае, если железо находится в двухвалентной форме, что обеспечивается аскорбиновой кислотой (витамин C). Дефицит аскорбиновой кислоты нарушает процесс гидроксилирования, что влияет на дальнейшие стадии синтеза коллагена: гликозилирование, отщепление N- и С-концевых пептидов и др. В результате синтезируется аномальный коллаген, более рыхлый. Эти изменения лежат в основе развития цинги. Коллаген и эластин формируют своеобразную «основу» кожи, которая предотвращает её обвисание, обеспечивает её эластичность и упругость. Эластин как белок прекращает выработку ферментов в человеческом организме в 14 лет, а коллаген — в 21—25, после чего кожные покровы не восстанавливаются и кожа стареет. Также важнейшим компонентом соединительной ткани является кератин — семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов биологического происхождения уступает лишь хитину. В основном из кератинов состоят роговые производные эпидермиса кожи — такие структуры, как волосы, ногти, рога, перья и др.
Фотография[править | править код]
Белок является основой для фотографической желатины, которая вместе с микрокристаллами галогенидов серебра образует фотографическую эмульсию. При получении фотографической желатины коллаген денатурируют кислотой или щёлочью. Фотографическая эмульсия, нанесённая тонким слоем на целлулоидную плёнку, стекло или бумагу, а затем высушенная, — это и есть светочувствительный слой фотоматериала (например, фотоплёнки).
В настоящее время описано 28 типов коллагена, которые кодируются более чем 40 генами. Они отличаются друг от друга по аминокислотной последовательности, а также по степени модификации — интенсивности гидроксилирования или гликозилирования. Общим для всех коллагенов является существование 1 или более доменов, содержащих тройную спираль и присутствие их во внеклеточном матриксе. Более 90 % всего коллагена высших организмов приходится на коллагены I, II,III и IV типов.
| Разновидности коллагена | Типы |
|---|---|
| Фибриллярные коллагены | I, II, III, V, XI, XXIV, XXVII |
| Фибрилл-ассоциированные коллагены (FACIT) | IX, XII, XIV, XVI, XIX, XX, XXI, XXII |
| Коллагены, формирующие филаменты- бусины (beaded filament forming) | VI |
| Сетеобразующие коллагены | IV, VIII, X |
| Коллаген, формирующий якорные фибриллы | VII |
| Трансмембранные коллагены | XIII, XVII, XXIII, XXV/CLAC-P |
| Другие коллагены | XXVIII, XV, XVIII |
Кроме коллагеновых белков существует множество других белков, содержащих в своей структуре домен с тройной коллагеновой спиралью[7][8]. И, тем не менее, их не причисляют к коллагенам, а только к «коллагеноподобным». К большой группе коллагеноподобных белков относятся подкомпонент C1q-комплемента, C1q-подобный фактор, адипонектин, колектины и фиколины, концевая структура ацетилхолинестаразы, три макрофаговых рецептора, эктодисплазин и EMILIN. Эти белки, так же как и коллагены, играют структурную и регуляторную роль.
Коллаген первого типа, самый архетипичный, является тримерным белком, собирающимся в тройные спирали без разрывов, самособирающимся в фибриллы и обладающим наибольшей механической прочностью. Между тем, все остальные коллагены отличаются от него в одном или нескольких аспектах. Некоторые коллагены имеют разрывы в тройной спирали и не обязательно собираются в фибриллы.
| Тип коллагена | Гены | Молекулы | Органы | Ассоциированные болезни |
|---|---|---|---|---|
| I | COL1A1 COL1A2 | α1(I)2α2(I), α1(I)3 | Повсеместно в мягких и твёрдых тканях, в коже, костях, роговице глаза, в склере, в стенке артерий и др. | Синдром Элерса-Данлоса, остеогенез, ревматизм, синдром Марфана, дисплазии |
| II | COL2A1 | α1(II)3 + см тип XI | Гиалиновые и фиброзные хрящи, стекловидное тело, роговица | Коллагенопатия II и XI типа, синдром Стиклера, ахондрогенез |
| III | COL3A1 | α1(III)3 | Дерма кожи плода, стенки крупных кровеносных сосудов, ретикулярные волокна органов кроветворения | Синдром Элерса-Данлоса, фибромышечная дисплазия, аневризма аорты |
| IV | COL4A1 COL4A2 COL4A3 COL4A4 COL4A5 COL4A6 | α1(IV)2α2(IV), другие непонятно | Базальные мембраны, капсула хрусталика | Синдром Альпорта, синдроме Гудпасчера |
| V | COL5A1 COL5A2 COL5A3 | α1(V)2α2(V), α1(V)α2(V)α3(V) + см тип XI | Мягкие ткани, плацента, сосуды, хорион | Синдром Элерса-Данлоса |
| VI | COL6A1 COL6A2 COL6A3 COL6A4 COL6A5 COL6A6 | α1(VI)α2(VI)α3(VI) | Микрофибриллы в мягких тканях и хрящах | Миопатия Ульриха, миопатия Бэтлема, атопический дерматит |
| VII | COL7A1 | α1(VII)3 | Якорные фибриллы в связке кожи и эпидермиса | Буллезный эпидермолиз |
| VIII | COL8A1 COL8A2 | α1(VIII)α2(VIII) | Роговица, эндотелий | Дистрофия роговицы |
| IX | COL9A1 COL9A2 COL9A3 | α1(IX)α2(IX)α3(IX) | Хрящи, стекловидное тело | Синдром Стиклера, остеоартрит, эпифизарная дисплазия |
| X | COL10A1 | α1(X)3 | Гипертрофическая зона области роста | Метафизарная дисплазия Шмида |
| XI | COL11A1 COL11A2 | α1(XI)α2(XI)α1(II), α1(XI)α2(V)α1(II) | Хрящи, стекловидное тело | Коллагенопатия II и XI типов, остеопороз |
| XII | COL12A1 | α1(XII)3 | Мягкие ткани | Повреждения сухожилий |
| XIII | COL13A1 | α1(XIII)3 | Поверхность клеток, эпителиальные клетки | |
| XIV | COL14A1 | α1(IV)3 | Мягкие ткани | |
| XV | COL15A1 | α1(XV)3 | Эндотелиальные клетки | Карцинома |
| XVI | COL16A1 | α1(XVI)3 | Повсеместно | |
| XVII | COL17A1 | α1(XVII)3 | Поверхность эпидермальных клеток | Буллезный эпидермиолиз, пузырчатка |
| XVIII | COL18A1 | α1(XVIII)3 | Эндотелиальные клетки | |
| XIX | COL19A1 | α1(XIX)3 | Повсеместно | Меланома, карцинома |
| XX | COL20A1 | α1(XX)3 | Выделен из куриного эмбриона | |
| XXI | COL21A1 | α1(XXI)3 | Кровеносные сосуды | |
| XXII | COL22A1 | α1(XXII)3 | Только в местах мышечно-сухожильных соединений | |
| XXIII | COL23A1 | α1(XXIII)3 | Опухолевые клетки | |
| XXIV | COL24A1 | α1(XXIV)3 | Формирующиеся кости | Остеохондроз |
| XXV | COL25A1 | α1(XXV)3 | Атеросклеротические бляшки | Болезнь Альцгеймера |
| XXVI | COL26A1=EMID2 | α1(XXVI)3 | Половые органы | |
| XXVII | COL27A1 | α1(XXVII)3 | Мягкие ткани | |
| XXVIII | COL28A1 | α1(XXVIII)3 | Нервная система |
Нарушения синтеза коллагена в организме лежат в основе таких наследственных заболеваний, как дерматоспораксис у животных, латиризм (характерна разболтанность суставов, привычные вывихи), синдром Элерса-Данлоса (до 14 типов проявлений), несовершенный остеогенез (болезнь «стеклянного человека», врождённый рахит, врождённая ломкость костей), болезнь Марфана, муковисцидоз.
Характерным проявлением этих заболеваний является повреждение связочного аппарата, хрящей, костной системы, наличие пороков сердечных клапанов.
Болезни, вызванные дефектами при биосинтезе коллагена, в том числе так называемые коллагенозы, возникают из-за множества причин. Это может быть из-за мутации в гене, кодирующем аминокислотную последовательность ферментов, продуцирующих коллаген, приводящей к изменению формы коллагеновой молекулы, или ошибки в посттранстляционной модификации коллагена. Также болезни могут быть вызваны недостатком или «неправильной работой» ферментов, вовлечённых в биосинтез коллагена — дефицит ферментов гидроксилирования (пролин-, лизингидроксилазы), гликозилтрансфераз, N-проколлагеновой и С-проколлагеновой пептидаз, лизилоксидаз с последующим нарушением поперечных сшивок, дефицит меди, витаминов B6, B13 (оротовая кислота), C. При приобретённых болезнях, таких как цинга, восстановление баланса ферментов до нормального может привести к полному излечению.
Практически любая генная мутация ведёт к утрате или изменению функций коллагена, что, в свою очередь, отражается на свойствах тканей и органов. Генные мутации в коллагеновом домене могут привести к изменению формы тройной спирали путём вставки/выпадения аминокислоты из полипептидной цепочки или замены Gly на другое основание. Мутации в неколлагеновых доменах могут привести к неправильной пространственной сборке α-цепей в надмолекулярные структуры (фибриллы или сети), что также ведёт к утрате функций. Мутантные α-цепи способны образовывать трёхспиральный комплекс с нормальными α-цепями. В большинстве случаев, такие комплексы нестабильны и быстро разрушаются, однако, такая молекула может и нормально выполнять свою роль, если не затронуты функционально важные области. Большинство болезней, вызванных мутациями в «коллагеновых» генах, являются доминантными.
- ↑ Fibrillar collagen: the key to vertebrate evolution? A tale of molecular incest. Boot-Handford RP, Tuckwell DS. Bioessays. 2003 Feb;25(2):142-51.
- ↑ Di Lullo, Gloria A.; Sweeney, Shawn M.; Körkkö, Jarmo; Ala-Kokko, Leena; San Antonio, James D. Mapping the Ligand-binding Sites and Disease-associated Mutations on the Most Abundant Protein in the Human, Type I Collagen (англ.) // Journal of Biological Chemistry|J. Biol. Chem. : journal. — 2002. — Vol. 277, no. 6. — P. 4223—4231. — DOI:10.1074/jbc.M110709200. — PMID 11704682.
- ↑ 1 2 Тропоколлаген (Tropocollagen) в словаре dic.academic.ru
- ↑ XuMuK.ru — КОЛЛАГЕН — Химическая энциклопедия
- ↑ Би-би-си | Наука и техника | Найдены мягкие ткани тираннозавра? — 25 марта 2005 г.
- ↑ Элементы — новости науки: Коллаген из костей динозавров — это уже реальность. — 20.04.07
- ↑ Franzke CW, Bruckner P, Bruckner-Tuderman L. Collagenous transmembrane proteins: recent insights into biology and pathology. J Biol Chem. 2005 Feb 11;280(6):4005-8.
- ↑ Hoppe HJ, Reid KB. Collectins—soluble proteins containing collagenous regions and lectin domains—and their roles in innate immunity. Protein Sci. 1994 Aug;3(8):1143-58
Коллаген. «БИОЛОГИЧЕСКАЯ ХИМИЯ», Березов Т.Т., Коровкин Б.Ф.
Коллаген
Характерным компонентом структуры соединительной ткани являются коллагеновые волокна. Они построены в основном из своеобразного белкаи – коллагена. Коллаген составляет 25–33% от общего количества белка организма взрослого человека, или 6% от массы тела.
Видимые в оптическом микроскопе коллагеновые волокна состоят из различимых в электронном микроскопе фибрилл – вытянутых в длину белковых молекул, названных тропоколлагеном. Тропоколлаген – основная структурная единица коллагена (рис. 21.2). Необходимо четко разграничивать понятия «коллагеновые волокна» и «коллаген». Первое понятие по существу является морфологическим и не может быть сведено к биохимическим представлениям о коллагене как о белке. Коллагеновое волокно представляет собой гетерогенное образование и содержит, кроме белка коллагена, другие химические компоненты. Молекула тропоколла-гена – это белок коллаген. Одной из отличительных черт данного белка является то, что 1/3 всех его аминокислотных остатков составляет глицин, 1/3 – пролин и 4-гидроксипролин, около 1% – гидроксилизин; некоторые молекулярные формы коллагена содержат также 3-гидроксипролин, хотя и в весьма ограниченном количестве:
Молекулярная масса тропоколлагена около 285000. Тропоколлаген состоит из трех полипептидных цепей одинакового размера, которые сливаются в спиралевидный триплет. Тройная спираль стабилизируется многочисленными межцепочечными поперечными сшивками между лизи-новыми и гидроксилизиновыми остатками. Каждая полипептидная цепь тропоколлагена содержит около 1000 аминокислотных остатков. Таким образом, основная структурная единица коллагена имеет очень большие размеры, например в 10 раз больше, чем химотрипсин.
Изучение аминокислотного состава и последовательности чередования аминокислот в полипептидных цепях тропоколлагена показало, что существует два типа цепей – цепи α1 и α2, а также четыре разновидности цепи α1: α1 (I), α1 (II), α1 (III) и α1 (IV). В табл. 21.1 представлены данные о структуре коллагенов различных тканей.
Как и все белки, коллаген синтезируется клетками из свободных аминокислотных остатков. Аминокислотные остатки, специфичные для молекулы коллагена, гидроксипролин и гидроксилизин не образуются из соответствующих свободных аминокислот. Эти аминокислотные остатки появляются после включения пролина и лизина в полипептидную цепь с участием ферментов пролилгидроксилазы или лизилгидроксилазы и кофактора – аскорбиновой кислоты.
Учитывая наличие разных молекулярных форм в пределах одного типа (например, коллаген типа 1 имеет состав [α1 (I)]2 α2 либо [α1 (I)]3, есть основание считать, что существует по крайней мере не менее 10 молекулярных форм коллагена (Е.С. Северин).
Напомним, что коллаген – внеклеточный белок, но он синтезируется в виде внутриклеточной молекулы-предшественника, которая перед образованием фибрилл зрелого коллагена подвергается посттрансляционной модификации. Предшественник коллагена (сначала препроколлаген, а затем проколлаген) претерпевает процессинг в ходе прохождения через эндо-плазматический ретикулум и комплекс Гольджи до появления во внеклеточном пространстве. Внеклеточные амино- и карбоксипротеаза проколлагена удаляют соответственно аминоконцевой и карбоксиконцевой про-пептиды. Вновь образованные молекулы коллагена спонтанно собираются в коллагеновые фибриллы. В результате перекрестного связывания цепей и спиральных молекул фибрилл через основания Шиффа и альдольную конденсацию (т.е. перекрестное связывание их рядом ковалентных связей) образовавшиеся фибриллы приобретают силу напряжения зрелых колла-геновых фибрилл.
Предыдущая страница | Следующая страница
СОДЕРЖАНИЕ
Еще по теме:Виды коллагена
Коллаген — полиморфный белок, в настоящее время известно 19 типов коллагена, которые отличаются друг от друга по первичной структуре пептидных цепей, функциям и локализации в организме. 95% всего коллагена в организме человека составляют коллагены I, II и III типов.
Типы | Гены | Ткани и органы |
I | COLIA1, COL1A2 | Кожа, сухожилия, кости, роговица, плацента, артерии, печень, дентин |
II | COL2A1 | Хрящи, межпозвоночные диски, стекловидное тело, роговица |
III | C0L3A1 | Артерии, матка, кожа плода, строма паренхиматозных органов |
IV | COL4A1-COL4A6 | Базальные мембраны |
V | COL5A1-COL5A3 | Минорный компонент тканей, содержащих коллаген I и II типов (кожа, роговица, кости, хрящи, межпозвоночные диски, плацента) |
VI | COL6A1-COL6A3 | Хрящи, кровеносные сосуды, связки, кожа, матка, лёгкие, почки |
VII | COL7A1 | Амнион, кожа, пищевод, роговица, хорион |
VIII | COL8A1-COL8A2 | Роговица, кровеносные сосуды, культуральная среда эндотелия |
IX | COL9A1-COL9A3 | Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
X | COL10A1 | Хрящи (гипертрофированные) |
XI | COLUA1-COL11A2 | Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
XII | COL12A1 | Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XIII | C0L13A1 | Многие ткани |
XIV | COL14A1 | Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XV | C0L15A1 | Многие ткани |
XVI | COL16A1 | Многие ткани |
XVII | COL17A1 | Гемидесмосомы кожи |
XVIII | COL18A1 | Многие ткани, например печень, почки |
XIX | COL19A1 | Клетки рабдомиосаркомы |
Гены коллагена называются по типам коллагена и записываются арабскими цифрами, например СОL1 — ген коллагена 1 типа, COL2 — ген коллагена II типа и т.д. К этому символу приписываются буква А (обозначает α-цепь) и арабская цифра (обозначает вид α-цепи). Например, COL1A1 и COL1A2 кодируют, соответственно, α1, и α2-цепи коллагена I типа.
Этапы синтеза и созревания коллагена
Синтез и созревание коллагена — сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе:
1. На полисомах ЭПР синтезируются полипептидные препро-α-цепиколлагена. Они содержит начиная сN-конца: 1). гидрофобный «сигнальный» пептид, содержащий около 100 АК; 2). N-концевой пропептид, содержащий около 100 АК, в том числе цистеин; 3). α-цепь коллагена 4). С-концевой пропептид, содержащий около 250 АК, в том числе цистеин. КонцевыеС- и N-пропептидыформируют глобулярные домены и необходимы для правильного формирования тройной спирали.«Сигнальный» пептид, обеспечивает поступление синтезируемой на рибосоме препро-α-цепи в полость ЭПР.
2. В полости ЭПР при отщеплении сигнального пептида препро-α-цепиколлагена превращаются впро-α-цепи.
3. Поступающие в полости ЭПР про-α-цепи коллагенаподвергаются модификации.
а). Цистеины N-пропептидов образуют внутрицепочечные дисульфидные мостики, формируя наN-конце глобулярную структуру;
б). Пролины и лизины в Y-положении (гли-х-у) про-α-цепей гидроксилируются пролил-4-гидроксилазойилизил-5-гидроксилазойв 4-гидроксипролины (Hyp) и 5-гидроксилизины (Hyl). Некоторые пролины в Х-положениях гидроксилируются в 3-гидроксипролины пролил-3-гидроксилазой. Оксигеназы (гидроксилазы), содержат Fe2+, находятся на мембране ЭПР. Для реакции необходимы а-КГ, О2и витамин С:
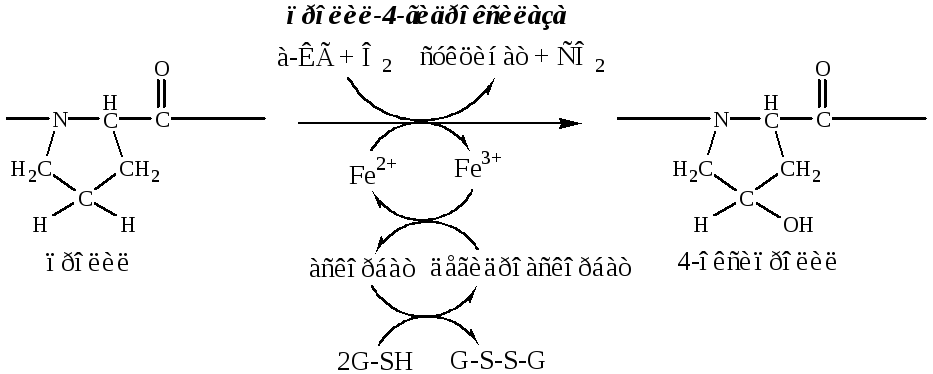
Гидроксилирование пролина необходимо для стабилизации тройной спирали коллагена, ОН-группы гидроксипролина участвуют в образовании водородных связей.
Гидроксилирование лизина необходимо для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл.
в). Гидроксилизин про-α-цепей при участии гликозилтрансфераз гликозилируется галактозой или галактозилглюкозой. В молекуле коллагена сухожилий (тип I) количество углеводов равно 6, а в моллекуле коллагена капсулы хрусталика (тип TV) — 110. Роль этих углеводных групп неясна.
4). В просвете ЭПР после отделения от рибосом про-α-цепей, 3 из них с помощью С-концевых пропептидов соединяются между собой дисульфидными мостиками (цистеины С-пропептидов образуют внутри- и межцепочечные дисульфидные мостики) и скручиваются с образованием тройной спирали проколлагена. Тройная спираль проколлагена стабилизируется водородными связями. После этого гидроксилирование и гликозилирование про-α-цепей прекращается.
5). Из ЭПР молекулы проколлагена перемещаются в аппарат Гольджи, включаются в секреторные пузырьки и секретируются в межклеточное пространство.
6). В межклеточном матриксе от некоторых проколлагенов(I, II, III, V, XI типов)проколлагенпептидазыотщепляют концевые С- иN-пропептиды, в результате чего образуетсятропоколлагены. У проколлагенов IV, VIII, X типов концевые пропептиды не отщепляются.
Синтезированные молекулы проколлагенов и тропоколлагенов способны образовывать различные структуры. Например, тропоколлагены образуют фибриллы, а проколлагены образуют сети.
Структура | Тип коллагена |
Фибриллы | I, II, III – основные. V, XI — минорные |
Ассоциированные с фибриллами | IX, XII, XIV, XVI, XIX |
Сети | IV, VIII, X |
Микрофибриллы | VI |
«Заякоренные» фибриллы | VII |
Трансмембранные домены | XIII, XVII |
Другие | XV, XVIII |

ТОП 10: |
Стр 1 из 4Следующая ⇒ Структура и свойства коллагена. Коллаге́н — фибриллярный белок, составляющий основу соединительной ткани организма (сухожилие, кость, хрящ, дерма и т. п.) и обеспечивающий её прочность и эластичность. Коллаген обнаружен у многоклеточных животных; отсутствует у растений, бактерий, вирусов, простейших и грибов. Это основной компонент соединительной ткани и самый распространённый белок у млекопитающих, составляющий от 25% до 35% белков во всём теле. Свойства. Продуктом денатурации коллагена является желатин. Температура денатурации макромолекулы коллагена близка к температуре фибриллогенеза. Это свойство молекулы коллагена делает её максимально чувствительной к мутационным заменам. Структура. Молекула коллагена представляет собой правозакрученную спираль из трёх α-цепей. Такое образование известно под названием тропоколлаген. Один виток спирали α-цепи содержит три аминокислотных остатка. Молекулярная масса коллагена около 300 кДа, длина 300 нм, толщина 1,5 нм. Для первичной структуры белка характерно высокое содержание глицина, низкое содержание серосодержащих аминокислот и отсутствие триптофана. Коллаген относится к тем немногим белкам животного происхождения, которые содержат остатки нестандартных аминокислот: около 21 % от общего числа остатков приходится на 3-гидроксипролин, 4-гидроксипролин и 5-гидроксилизин[4]. Каждая из α-цепей состоит из триад аминокислот. В триадах третья аминокислота всегда глицин, вторая — пролин или лизин, первая — любая другая аминокислота, кроме трёх перечисленных[3]. Коллаген существует в нескольких формах. Основная структура всех типов коллагена является схожей. Коллагеновые волокна образуются путём агрегации микрофибрилл, имеют розовый цвет при окраске гематоксилином и эозином и голубой или зелёный при различных треххромных окрасках, при импрегнации серебром окрашиваются в буро-жёлтый цвет. Фибриллярная структура Тропоколлагены (структурные единицы коллагена) спонтанно объединяются, прикрепляясь друг к другу смещенными на определённое расстояние концами, образуя в межклеточном веществе более крупные структуры. В фибриллярных коллагенах молекулы смещены относительно друг друга примерно на 67нм (единица, которая обозначается буквой «D» и меняется в зависимости от состояния гидратации вещества). В целом каждый D-период содержит четыре целых и часть пятой молекулы коллагена. Величина 300 нм, поделенная на 67 нм (300:67) не дают целого числа и длина молекулы коллагена разделена на непостоянные по величине отрезки D. Следовательно, в разрезе каждого повтора D-периода микрофибриллы есть часть, состоящая из пяти молекул, называемая «перекрытие», и часть, состоящая из четырёх молекул — «разрыв». Тропоколлагены к тому же скомпонованы в шестиугольную или псевдошестиугольную (в поперечном разрезе) конструкцию, в каждой области «перекрытия» и «разрыва». Внутри тропоколлагенов существует ковалентная связь между цепями, а также некоторое непостоянное количество данных связей между самими тропоколагеновыми спиралями, образующими хорошо организованные структуры (например, фибриллы). Более толстые пучки фибрилл формируются с помощью белков нескольких других классов, включая другие типы коллагенов, гликопротеины, протеогликаны, использующихся для формирования различных типов тканей из разных комбинаций одних и тех же основных белков. Нерастворимость коллагена была препятствием к изучению мономера коллагена, до того момента как было обнаружено, что возможно извлечь тропоколлаген молодого животного, поскольку он ещё не образовал сильных связей с другими субъединицами фибриллы. Тем не менее, усовершенствование микроскопов и рентгеновских аппаратов облегчили исследования, появлялось все больше подробных изображений структуры молекулы коллагена. Эти поздние открытия очень важны для лучшего понимания того, как структура коллагена влияет на связи между клетками и межклеточным веществом, как ткани меняются во время роста и регенерации, как они меняются во время эмбрионального развития и при патологии. Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена. Коллагеновые волокна — это пучки фибрилл.
2.Цинга. Процессы гидроксилированияпролина и лизина, их роль в возникновении цинги. Цинга́ — болезнь, вызываемая острым недостатком витамина C (аскорбиновая кислота), который приводит к нарушению синтеза коллагена, и соединительная ткань теряет свою прочность. Гидроксилированиепролина и лизина начинается в период трансляции коллагеновой мРНК на рибосомах и продолжается на растущей полипептидной цепи вплоть до её отделения от рибосом. После образования тройной спирали дальнейшее гидроксилированиепролиловых и лизиловых остатков прекращается. Реакции гидроксилирования катализируют ок-сигеназы, связанные с мембранами микросом. Пролиловые и лизиловые остатки в Y-положении пептида (Гли-х-у)n подвергаются действию, соответственно, пролил-4-гидроксилазы и лизил-5-гидроксилазы. Пролил-3-гидроксилаза действует на некоторые остатки пролина в Х-положениях. Необходимыми компонентами этой реакции являются оскетоглутарат, О2 и витамин С (аскорбиновая кислота). Донором атома кислорода, который присоединяется к С-4 пролина, является молекула О2, второй атом О2 включается в сукцинат, который образуется при декарбоксилировании α-кетоглутарата, а из карбоксильной группы а-кетоглутарата образуется СО2 (см. схему А на с. 691). Гидроксилазыпролина и лизина содержат в активном центре атом железа Fe2+. Для сохранения атома железа в ферроформе необходим восстанавливающий агент. Роль этого агента выполняет кофермент гидроксилаз — аскорбиновая кислота, которая легко окисляется в дегидроаскорбиновую кислоту. Обратное превращение происходит в ферментативном процессе за счёт восстановленного глутатиона (см. схему Б на с. 691). Гидроксилированиепролина необходимо для стабилизации тройной спирали коллагена, ОН-группыгидроксипролина (Hyp) участвуют в образовании водородных связей. А гидроксилирование лизина очень важно для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл. При цинге — заболевании, вызванном недостатком витамина С, нарушается гидроксилирование остатков пролина и лизина. В результате этого образуются менее прочные и стабильные коллагеновые волокна, что приводит к большой хрупкости и ломкости кровеносных сосудов с развитием цинги. Клиническая картина цинги характеризуется возникновением множественных точечных кровоизлияний под кожу и слизистые оболочки, кровоточивостью дёсен, выпадением зубов, анемией.
Биохимия мукополисахаридов. Мукополисахариды, полимерные углевод-белковые комплексы с преимущественным содержанием углеводной части (70-80%). Наиболее изучены кислые Мукополисахариды различных видов соединительной ткани и некоторых жидкостей организма (синовиальная жидкость суставов, стекловидное тело глаза). Основные представители Мукополисахариды: гиалуроновая кислота, гепарин, хондроитинсерные кислоты, кератосульфат (входит в состав хрящей и роговицы глаза). Углеводная часть кислых Мукополисахариды — линейный полисахарид с периодически повторяющимся звеном, состоящим из остатка N-сульфо- или N-ацетиламиносахара (D-глюкозамина или D-галактозамина) и уроновой кислоты. Остатки серной кислоты в составе сульфатированных Мукополисахариды связаны с гидроксильными группами моносахаридных компонентов. Кислые Мукополисахариды сильно различаются по молекулярной массе, прочности связывания компонентов и по функциональным свойствам. Благодаря способности связывать и удерживать воду кислые Мукополисахариды служат природным смазочным материалом суставов и определяют эластичность соединительной ткани; входя в состав хрящей и связок, Мукополисахариды выполняют опорно-двигательные функции. Мукополисахариды обладают бактерицидными свойствами. Состав Мукополисахариды соединительной ткани меняется при старении. Нарушения обмена Мукополисахариды вызывают изменение состава соединительной ткани и жидкостей организма и приводят к ряду заболеваний (коллагенозы, мукополисахаридозы, ревматизм и др.).
Химический состав слюны. Слюна обладает pH от 5,6 до 7,6. На 98,5 % и более состоит из воды, содержит соли различных кислот, микроэлементы и катионы некоторых щелочных металлов, лизоцим и другие ферменты, некоторые витамины. Основными органическими веществами слюны являются белки, синтезируемые в слюнных железах (некоторые ферменты, гликопротеиды, муцины, иммуноглобулины класса А) и вне их. Часть белков слюны имеет сывороточное происхождение (некоторые ферменты, альбумины, β-липопротеиды, иммуноглобулины классов G и М и др.). Химический состав слюны подвержен суточным колебаниям, он также зависит от возраста (у пожилых людей, например, значительно повышается количество кальция, что имеет значение для образования зубного и слюнного камня). Изменения в составе слюны могут быть связаны с приемом лекарственных веществ и интоксикациями. Состав слюны меняется также при ряде патологических состояний и заболеваний. Так, при обезвоживании организма происходит резкое снижение слюноотделения; при сахарном диабете в слюне увеличивается количество глюкозы; при уремии в слюне значительно возрастает содержание остаточного азота. Уменьшение слюноотделения и изменения в составе слюны приводят к нарушениям пищеварения, заболеваниям зубов.
11-12. основные черты механизма секреции слюны. Функции слюнных желез. Определение скорости секреции слюны. Возрастные изменения скорости секреции слюны. . Регуляция слюноотделения преимущественно осуществляется нервными механизмами. Вне пищеварения в основном функционируют мелкие железы. В пищеварительный период секреция слюны значительно возрастает. Регуляция пищеварительной секреции осуществляется условно – и безусловнорефлекторными механизмами. Безусловнорефлекторное слюноотделение возникает при раздражении первоначально тактильных, а затем температурных и вкусовых рецепторов полости рта. Но основную роль играют вкусовые. Нервные импульсы от них по афферентным нервным волокнам язычного, языкоглоточного и верхнегортанного нервов поступают в слюноотделительный центр продолговатого мозга. Он находится в области ядер лицевого и языкоглоточного нервов. От центра импульсы по эфферентным нервам идут к слюнным железам. К околоушной железе эффернтные парасимпатические волокна идут от нижнего слюноотделительного ядра в составе нерва Якобсона, а затем ушно-височных нервов. Парасимпатические нервы, иннервирующие серозные клетки подчелюстных и подъязычных желез начинаются от верхнего слюноотделительного ядра, идут в составе лицевого нерва, а затем барабанной струны. Симпатические нервы иннервирующие железы идут от слюннотделительных ядер II – VI грудных сегментов, прерываются в шейном ганглии, а затем их постганглионарные волокна идут к слизистым клеткам. Поэтому раздражение парасимпатических нервов ведет к выделению большого количества жидкой слюны, а симпатических – небольшого объема слизистой. Условно-рефлекторное слюноотделение начинается раньше безусловно рефлекторного. Оно возникает на запах, вид пищи, звуки предшествующие кормлению. Условно-рефлекторные механизмы секреции обеспечиваются корой больших полушарий, которая через нисходящие пути стимулирует центр слюноотделения. Функции слюнных желез.экзокринная — секреция белковых и слизистых компонентов слюны; эндокринная — секреция гормоноподобных веществ; фильтрационная — фильтрация жидкостных компонентов плазмы крови из капилляров в состав слюны; экскреторная — выделение конечных продуктов метаболизма.
13. Структура и формирование кости.Все кости скелета человека образованы пористой тканью, покрытой твердым материалом, преимущественно кальцием и фосфором, которые и придают костям нужную форму и обеспечивают их прочность.Кости и зубы содержат более 90 % кальция, имеющегося в организме человека. Каждая кость скелета — это живая, активно функционирующая и непрерывно обновляющаяся структура.Для сохранения твердости кости нуждаются в регулярной нагрузке, а в случае её недостатка, кости подвергаются патологическим изменениям («кальций вымывается из кости»). Этот быстротекущий процесс можно наблюдать, например, на загипсованной в течение месяца ноге.Содержание кальция в костях уменьшается и с возрастом, при этом отмечается их хрупкость. У пожилых людей даже при незначительных травмах и ушибах часто случаются переломы костей. Недостаток кальция, и как следствие этого остеопороз, возникают почти у каждого человека в связи со старением организма и поэтому могут рассматриваться как естественное явление.К декальцинации организма в целом и костей скелета в частности могут привести ряд заболеваний кишечника, связанных с пониженной способностью организма получать кальций из пищи; заболевания почек, выделяющих ненормально большое количество кальция. Употребление некоторых медикаментозных препаратов, например, гормональных, также влияет на содержание кальция в костях и может способствовать его излишнему выведению.Кроме структурно-опорной функции (в составе костной ткани)кальций участвует в проведении нервного импульса, в нервно-мышечном сокращении, в работе системы свёртывания крови, тканевом дыхании, активирует ряд ферментов, обладает десенсибилизирующим (антигистаминным) действием.Необходим кальций и для восстановления клеток всего организма. Если в рационе питания отсутствует органический кальций, то от этого будут страдать не только кости, но и другие части тела.В естественных минералах нуждаются не только позвоночник и кости, но и спинной мозг. Наиболее характерным признаком недостатка кальция в крови является повышенная нервная возбудимость(нервные волокна не проводят соответствующие сигналы, тело не расслабляется). У детей такое состояние сопровождается капризным поведением, вспышками раздражения, могут появиться мышечные судороги, спазмы.Суточная потребность в кальции — 800 — 1100 мг.Больше всего кальция содержится в молоке и молочных продуктах, а также в овощах с зелёными листьями, в кочанной капусте, черносливе, крапиве, красном перце, петрушке, мяте перечной, подорожнике, в блюдах из овсянки и плодах шиповника.
14. Факторы, влияющие на метаболизм кости. К факторам, влияющим на метаболизм костной ткани, прежде всего следует отнести гормоны, ферменты и витамины. Многие аспекты данной проблемы уже рассматривались в предыдущих главах. В данном разделе будут приведены лишь краткие сведения.Известно, что минеральные компоненты костной ткани находятся практически в состоянии химического равновесия с ионами кальция и фосфата сыворотки крови. Поступление, депонирование и выделение кальция и фосфата регулируются весьма сложной системой, в которой среди других факторов важная роль принадлежит паратгормону (гормон околощитовидных желез) и кальцитонину (гормон щитовидной железы). При уменьшении концентрации ионов Са2+ в сыворотке крови возрастает секреция паратгормона (см. гл. 8). Непосредственно под влиянием этого гормона в костной ткани активируются клеточные системы, участвующие в резорбции кости (увеличение числа остеокластов и их метаболической активности), т.е. остеокласты способствуют повышенному растворению содержащихся в костях минеральных соединений. Заметим, что паратгор-мон увеличивает также реабсорбцию ионов Са2+ в почечных канальцах. Суммарный эффект проявляется в повышении уровня кальция в сыворотке крови.В свою очередь при увеличении содержания ионов Са2+ в сыворотке крови секретируется гормон кальцитонин, действие которого состоит в снижении концентрации ионов Са2+ за счет отложения его в костной ткани. Иными словами, кальцитонин повышает минерализацию кости и уменьшает число остеокластов в зоне действия, т.е. угнетает процесс костной резорбции. Все это увеличивает скорость формирования кости.В регуляции содержания ионов Са2+ важная роль принадлежит витамину D, который участвует в биосинтезе Са2+-связывающих белков. Эти белки необходимы для всасывания ионов Са2+ в кишечнике, реабсорбции их в почках и мобилизации кальция из костей. Поступление в организм оптимальных количеств витамина D является необходимым условием для нормального течения процессов кальцификации костной ткани. При недостаточности витамина D эти процессы нарушаются. Прием в течение длительного времени избыточных количеств витамина D приводит к деминерализации костей.На развитие кости влияет также витамин А. Прекращение роста костей является ранним проявлением недостаточности витамина А. Считают, что данный факт обусловлен нарушением синтеза хондроитинсуль-фата. Показано также, что при введении животным высоких доз витамина А, превышающих физиологическую потребность и вызывающих развитие гипервитаминоза А, наблюдается резорбция кости, что может приводить к переломам.Для нормального развития костной ткани необходим витамин С. Действие витамина С не метаболизм костной ткани обусловлено прежде всего влиянием на процессе биосинтеза коллагена. Аскорбиновая кислота необходима для осуществления реакции гидроксилированияпролина и лизина. При недостаточности витамина С остеобласты не синтезируют «нормальный» коллаген, что приводит к нарушениям процессов обызвествления костной ткани. Недостаток витамина С вызывает также изменения в синтезе гликозаминогликанов: содержание гиалуроновой кислоты в костной ткани увеличивается в несколько раз, тогда как биосинтез хондроитин-сульфатов замедляется.
Структура и свойства коллагена. Коллаге́н — фибриллярный белок, составляющий основу соединительной ткани организма (сухожилие, кость, хрящ, дерма и т. п.) и обеспечивающий её прочность и эластичность. Коллаген обнаружен у многоклеточных животных; отсутствует у растений, бактерий, вирусов, простейших и грибов. Это основной компонент соединительной ткани и самый распространённый белок у млекопитающих, составляющий от 25% до 35% белков во всём теле. Свойства. Продуктом денатурации коллагена является желатин. Температура денатурации макромолекулы коллагена близка к температуре фибриллогенеза. Это свойство молекулы коллагена делает её максимально чувствительной к мутационным заменам. Структура. Молекула коллагена представляет собой правозакрученную спираль из трёх α-цепей. Такое образование известно под названием тропоколлаген. Один виток спирали α-цепи содержит три аминокислотных остатка. Молекулярная масса коллагена около 300 кДа, длина 300 нм, толщина 1,5 нм. Для первичной структуры белка характерно высокое содержание глицина, низкое содержание серосодержащих аминокислот и отсутствие триптофана. Коллаген относится к тем немногим белкам животного происхождения, которые содержат остатки нестандартных аминокислот: около 21 % от общего числа остатков приходится на 3-гидроксипролин, 4-гидроксипролин и 5-гидроксилизин[4]. Каждая из α-цепей состоит из триад аминокислот. В триадах третья аминокислота всегда глицин, вторая — пролин или лизин, первая — любая другая аминокислота, кроме трёх перечисленных[3]. Коллаген существует в нескольких формах. Основная структура всех типов коллагена является схожей. Коллагеновые волокна образуются путём агрегации микрофибрилл, имеют розовый цвет при окраске гематоксилином и эозином и голубой или зелёный при различных треххромных окрасках, при импрегнации серебром окрашиваются в буро-жёлтый цвет. Фибриллярная структура Тропоколлагены (структурные единицы коллагена) спонтанно объединяются, прикрепляясь друг к другу смещенными на определённое расстояние концами, образуя в межклеточном веществе более крупные структуры. В фибриллярных коллагенах молекулы смещены относительно друг друга примерно на 67нм (единица, которая обозначается буквой «D» и меняется в зависимости от состояния гидратации вещества). В целом каждый D-период содержит четыре целых и часть пятой молекулы коллагена. Величина 300 нм, поделенная на 67 нм (300:67) не дают целого числа и длина молекулы коллагена разделена на непостоянные по величине отрезки D. Следовательно, в разрезе каждого повтора D-периода микрофибриллы есть часть, состоящая из пяти молекул, называемая «перекрытие», и часть, состоящая из четырёх молекул — «разрыв». Тропоколлагены к тому же скомпонованы в шестиугольную или псевдошестиугольную (в поперечном разрезе) конструкцию, в каждой области «перекрытия» и «разрыва». Внутри тропоколлагенов существует ковалентная связь между цепями, а также некоторое непостоянное количество данных связей между самими тропоколагеновыми спиралями, образующими хорошо организованные структуры (например, фибриллы). Более толстые пучки фибрилл формируются с помощью белков нескольких других классов, включая другие типы коллагенов, гликопротеины, протеогликаны, использующихся для формирования различных типов тканей из разных комбинаций одних и тех же основных белков. Нерастворимость коллагена была препятствием к изучению мономера коллагена, до того момента как было обнаружено, что возможно извлечь тропоколлаген молодого животного, поскольку он ещё не образовал сильных связей с другими субъединицами фибриллы. Тем не менее, усовершенствование микроскопов и рентгеновских аппаратов облегчили исследования, появлялось все больше подробных изображений структуры молекулы коллагена. Эти поздние открытия очень важны для лучшего понимания того, как структура коллагена влияет на связи между клетками и межклеточным веществом, как ткани меняются во время роста и регенерации, как они меняются во время эмбрионального развития и при патологии. Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена. Коллагеновые волокна — это пучки фибрилл.
2.Цинга. Процессы гидроксилированияпролина и лизина, их роль в возникновении цинги. Цинга́ — болезнь, вызываемая острым недостатком витамина C (аскорбиновая кислота), который приводит к нарушению синтеза коллагена, и соединительная ткань теряет свою прочность. Гидроксилированиепролина и лизина начинается в период трансляции коллагеновой мРНК на рибосомах и продолжается на растущей полипептидной цепи вплоть до её отделения от рибосом. После образования тройной спирали дальнейшее гидроксилированиепролиловых и лизиловых остатков прекращается. Реакции гидроксилирования катализируют ок-сигеназы, связанные с мембранами микросом. Пролиловые и лизиловые остатки в Y-положении пептида (Гли-х-у)n подвергаются действию, соответственно, пролил-4-гидроксилазы и лизил-5-гидроксилазы. Пролил-3-гидроксилаза действует на некоторые остатки пролина в Х-положениях. Необходимыми компонентами этой реакции являются оскетоглутарат, О2 и витамин С (аскорбиновая кислота). Донором атома кислорода, который присоединяется к С-4 пролина, является молекула О2, второй атом О2 включается в сукцинат, который образуется при декарбоксилировании α-кетоглутарата, а из карбоксильной группы а-кетоглутарата образуется СО2 (см. схему А на с. 691). Гидроксилазыпролина и лизина содержат в активном центре атом железа Fe2+. Для сохранения атома железа в ферроформе необходим восстанавливающий агент. Роль этого агента выполняет кофермент гидроксилаз — аскорбиновая кислота, которая легко окисляется в дегидроаскорбиновую кислоту. Обратное превращение происходит в ферментативном процессе за счёт восстановленного глутатиона (см. схему Б на с. 691). Гидроксилированиепролина необходимо для стабилизации тройной спирали коллагена, ОН-группыгидроксипролина (Hyp) участвуют в образовании водородных связей. А гидроксилирование лизина очень важно для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл. При цинге — заболевании, вызванном недостатком витамина С, нарушается гидроксилирование остатков пролина и лизина. В результате этого образуются менее прочные и стабильные коллагеновые волокна, что приводит к большой хрупкости и ломкости кровеносных сосудов с развитием цинги. Клиническая картина цинги характеризуется возникновением множественных точечных кровоизлияний под кожу и слизистые оболочки, кровоточивостью дёсен, выпадением зубов, анемией.
|
Коллаген — Википедия
Коллаген — фибриллярный белок, составляющий основу соединительной ткани организма (сухожилие, кость, хрящ, дерма и т. п.) и обеспечивающий её прочность и эластичность. Коллаген обнаружен у многоклеточных животных; отсутствует у растений, бактерий, вирусов, простейших и грибов[1]. Коллаген — основной компонент соединительной ткани и самый распространённый белок у млекопитающих[2], составляющий от 25 % до 45% белков во всём теле.
История исследования
Учёные десятилетиями не могли понять молекулярное строение коллагена. Первое доказательство того, что коллаген имеет постоянное строение на молекулярном уровне, было представлено в середине 1930-х годов. С того времени много выдающихся учёных, включая Нобелевских лауреатов, таких как Фрэнсис Крик, Лайнус Полинг, Александр Рич, Ада Йонат, Хелен Берман, Вилеайнур Рамачандран, работали над строением мономера коллагена.
Несколько противоречащих друг другу моделей (несмотря на известное строение каждой отдельной пептидной цепи) дали дорогу для создания троично-спиральной модели, объяснившей четвертичное строение молекулы коллагена.
Видео по теме
Свойства
Продуктом денатурации коллагена является желатин. Температура денатурации макромолекулы коллагена близка к температуре фибриллогенеза. Это свойство молекулы коллагена делает её максимально чувствительной к мутационным заменам.
Структура
Молекула коллагена представляет собой левозакрученную спираль из трёх α-цепей. Такое образование известно под названием тропоколлаген[3]. Один виток спирали α-цепи содержит три аминокислотных остатка. Молекулярная масса коллагена около 300 кДа, длина 300 нм, толщина 1,5 нм.
Для первичной структуры белка характерно высокое содержание глицина, низкое содержание серосодержащих аминокислот и отсутствие триптофана. Коллаген относится к тем немногим белкам животного происхождения, которые содержат остатки нестандартных аминокислот: около 21 % от общего числа остатков приходится на 3-гидроксипролин, 4-гидроксипролин и 5-гидроксилизин[4]. Каждая из α-цепей состоит из триад аминокислот. В триадах третья аминокислота всегда глицин, вторая — пролин или лизин, первая — любая другая аминокислота, кроме трёх перечисленных[3].
Коллаген существует в нескольких формах. Основа строения всех видов коллагена является схожей. Коллагеновые волокна образуются путём агрегации микрофибрилл, имеют розовый цвет при окраске гематоксилином и эозином и голубой или зелёный при различных трёххромных окрасках, при импрегнации серебром окрашиваются в буро-жёлтый цвет.
Фибриллярная структура
Тропоколлагены (структурные единицы коллагена) спонтанно объединяются, прикрепляясь друг к другу смещенными на определённое расстояние концами, образуя в межклеточном веществе более крупные структуры. В фибриллярных коллагенах молекулы смещены относительно друг друга примерно на 67нм (единица, которая обозначается буквой «D» и меняется в зависимости от состояния гидратации вещества). В целом каждый D-период содержит четыре целых и часть пятой молекулы коллагена. Величина 300 нм, поделенная на 67 нм (300:67) не дают целого числа и длина молекулы коллагена разделена на непостоянные по величине отрезки D. Следовательно, в разрезе каждого повтора D-периода микрофибриллы есть часть, состоящая из пяти молекул, называемая «перекрытие», и часть, состоящая из четырёх молекул — «разрыв». Тропоколлагены к тому же скомпонованы в шестиугольную или псевдошестиугольную (в поперечном разрезе) конструкцию, в каждой области «перекрытия» и «разрыва».
Внутри тропоколлагенов существует ковалентная связь между цепями, а также некоторое непостоянное количество данных связей между самими тропоколагеновыми спиралями, образующими хорошо организованные структуры (например, фибриллы). Более толстые пучки фибрилл формируются с помощью белков нескольких других классов, включая другие типы коллагенов, гликопротеины, протеогликаны, использующихся для формирования различных типов тканей из разных комбинаций одних и тех же основных белков. Нерастворимость коллагена была препятствием к изучению мономера коллагена, до того момента как было обнаружено, что возможно извлечь тропоколлаген молодого животного, поскольку он ещё не образовал сильных связей с другими субъединицами фибриллы. Тем не менее, усовершенствование микроскопов и рентгеновских аппаратов облегчили исследования, появлялось все больше подробных изображений структуры молекулы коллагена. Эти поздние открытия очень важны для лучшего понимания того, как структура коллагена влияет на связи между клетками и межклеточным веществом, как ткани меняются во время роста и регенерации, как они меняются во время эмбрионального развития и при патологии.
Коллагеновая фибрилла — это полукристаллическая структурная единица коллагена. Коллагеновые волокна — это пучки фибрилл.
Использование
Пищевая промышленность
С точки зрения питания, коллаген и желатин являются белками низкого качества, так как они не содержат всех незаменимых аминокислот, необходимых человеку — это неполноценные белки. Производители основанных на коллагене пищевых добавок утверждают, что их продукты могут улучшить качество кожи и ногтей, а также здоровье суставов[источник не указан 3056 дней].
Относительно дешёвые, часто предлагаемые сегодня на рынке под видом источника свободных аминокислот гидролизаты коллагена не всегда способны удовлетворить потребности человека в свободных аминокислотах, так как эти продукты не содержат готовые к усвоению аминокислоты, а являются лишь частично «переваренными» экстрактами суставных тканей млекопитающих, птиц или обитателей моря. Например, гидролизаты коллагена почти полностью лишены аминокислоты L-глютамина, не отличающейся стойкостью к термическому воздействию и долгому хранению сырья, большая часть глютамина разрушается уже на первых этапах хранения и переработки сырья, имеющийся небольшой остаток практически полностью распадается во время термической экстракции хрящевой ткани.
Наиболее качественными источниками аминокислот являются препараты, содержащие так называемые «свободные аминокислоты». Так как именно свободные аминокислоты являются практически готовыми к усвоению, организму не нужно тратить пищеварительные ферменты, время и энергию на их переваривание. Они способны в кратчайшие сроки поступить в кровь и, будучи доставленными ею к местам, нуждающимся в дополнительном синтезе коллагена, тут же включаются в его формирование[источник не указан 2792 дня].
Косметические средства
Коллаген входит в состав косметических средств для :
- Образования воздухопроницаемого, влагоудерживающего слоя на поверхности кожи, обладающего пластифицирующими (разглаживающими) свойствами, со свойствами влажного компресса;
- Продления действия экстрактов, масел и др. в составе косметических композиций;
- Придания блеска волосам, создания коллагенового (защитного) слоя на поверхности волос.
Научные исследования
В 2005 году учёным удалось выделить коллаген из сохранившихся мягких тканей тираннозавра[5][нет в источнике] и использовать его химический состав как ещё одно доказательство родства динозавров с современными птицами[6].
Научные исследования в медицине
Синтез коллагена — сложный ферментативный многостадийный процесс, который должен быть обеспечен достаточным количеством витаминов и минеральных элементов. Синтез протекает в фибробласте и ряд стадий вне фибробласта. Важный момент в синтезе — реакции гидроксилирования, которые открывают путь дальнейшим модификациям, необходимым для созревания коллагена. Катализируют реакции гидроксилирования специфические ферменты. Так, образование 4-оксипролина катализирует пролингидроксилаза, в активном центре которой находится железо. Фермент активен в том случае, если железо находится в двухвалентной форме, что обеспечивается аскорбиновой кислотой (витамин С). Дефицит аскорбиновой кислоты нарушает процесс гидроксилирования, что влияет на дальнейшие стадии синтеза коллагена: гликозилирование, отщепление N- и С-концевых пептидов и др. В результате синтезируется аномальный коллаген, более рыхлый. Эти изменения лежат в основе развития цинги. Коллаген и эластин формируют своеобразную «основу» кожи, которая предотвращает её обвисание, обеспечивает её эластичность и упругость. Эластин как белок прекращает выработку ферментов в человеческом организме в 14 лет, а коллаген — в 21—25, после чего кожные покровы не восстанавливаются и кожа стареет. Также важнейшим компонентом соединительной ткани является кератин — семейство фибриллярных белков, обладающих механической прочностью, которая среди материалов биологического происхождения уступает лишь хитину. В основном из кератинов состоят роговые производные эпидермиса кожи — такие структуры, как волосы, ногти, рога, перья и др.
Фотография
Белок является основой для фотографической желатины, которая вместе с микрокристаллами галогенидов серебра образует фотографическую эмульсию. При получении фотографической желатины коллаген денатурируют кислотой или щёлочью. Фотографическая эмульсия, нанесённая тонким слоем на целлулоидную плёнку, стекло или бумагу, а затем высушенная, — это и есть светочувствительный слой фотоматериала (например, фотоплёнки).
Типы коллагена
В настоящее время описано 28 типов коллагена, которые кодируются более чем 40 генами. Они отличаются друг от друга по аминокислотной последовательности, а также по степени модификации — интенсивности гидроксилирования или гликозилирования. Общим для всех коллагенов является существование 1 или более доменов, содержащих тройную спираль и присутствие их во внеклеточном матриксе. Более 90 % всего коллагена высших организмов приходится на коллагены I, II,III и IV типов.
| Разновидности коллагена | Типы |
|---|---|
| Фибриллярные коллагены | I, II, III, V, XI, XXIV, XXVII |
| Фибрилл-ассоциированные коллагены (FACIT) | IX, XII, XIV, XVI, XIX, XX, XXI, XXII |
| Коллагены, формирующие филаменты- бусины (beaded filament forming) | VI |
| Сетеобразующие коллагены | IV, VIII, X |
| Коллаген, формирующий якорные фибриллы | VII |
| Трансмембранные коллагены | XIII, XVII, XXIII, XXV/CLAC-P |
| Другие коллагены | XXVIII, XV, XVIII |
Кроме коллагеновых белков существует множество других белков, содержащих в своей структуре домен с тройной коллагеновой спиралью[7][8]. И, тем не менее, их не причисляют к коллагенам, а только к «коллагеноподобным». К большой группе коллагеноподобных белков относятся подкомпонент C1q-комплемента, C1q-подобный фактор, адипонектин, колектины и фиколины, концевая структура ацетилхолинестаразы, три макрофаговых рецептора, эктодисплазин и EMILIN. Эти белки, так же как и коллагены, играют структурную и регуляторную роль.
Коллаген первого типа, самый архетипичный, является тримерным белком, собирающимся в тройные спирали без разрывов, самособирающимся в фибриллы и обладающим наибольшей механической прочностью. Между тем, все остальные коллагены отличаются от него в одном или нескольких аспектах. Некоторые коллагены имеют разрывы в тройной спирали и не обязательно собираются в фибриллы.
| Тип коллагена | Гены | Молекулы | Органы | Ассоциированные болезни |
|---|---|---|---|---|
| I | COL1A1 COL1A2 | α1(I)2α2(I), α1(I)3 | Повсеместно в мягких и твёрдых тканях, в коже, костях, роговице глаза, в склере, в стенке артерий и др. | Синдром Элерса-Данлоса, остеогенез, ревматизм, синдром Марфана, дисплазии |
| II | COL2A1 | α1(II)3 + см тип XI | Гиалиновые и фиброзные хрящи, стекловидное тело, роговица | Коллагенопатия II и XI типа, синдром Стиклера, ахондрогенез |
| III | COL3A1 | α1(III)3 | Дерма кожи плода, стенки крупных кровеносных сосудов, ретикулярные волокна органов кроветворения | Синдром Элерса-Данлоса, фибромышечная дисплазия, аневризма аорты |
| IV | COL4A1 COL4A2 COL4A3 COL4A4 COL4A5 COL4A6 | α1(IV)2α2(IV), другие непонятно | Базальные мембраны, капсула хрусталика | Синдром Альпорта, синдроме Гудпасчера |
| V | COL5A1 COL5A2 COL5A3 | α1(V)2α2(V), α1(V)α2(V)α3(V) + см тип XI | Мягкие ткани, плацента, сосуды, хорион | Синдром Элерса-Данлоса |
| VI | COL6A1 COL6A2 COL6A3 COL6A4 COL6A5 COL6A6 | α1(VI)α2(VI)α3(VI) | Микрофибриллы в мягких тканях и хрящах | Миопатия Ульриха, миопатия Бэтлема, атопический дерматит |
| VII | COL7A1 | α1(VII)3 | Якорные фибриллы в связке кожи и эпидермиса | Буллезный эпидермолиз |
| VIII | COL8A1 COL8A2 | α1(VIII)α2(VIII) | Роговица, эндотелий | Дистрофия роговицы |
| IX | COL9A1 COL9A2 COL9A3 | α1(IX)α2(IX)α3(IX) | Хрящи, стекловидное тело | Синдром Стиклера, остеоартрит, эпифизарная дисплазия |
| X | COL10A1 | α1(X)3 | Гипертрофическая зона области роста | Метафизарная дисплазия Шмида |
| XI | COL11A1 COL11A2 | α1(XI)α2(XI)α1(II), α1(XI)α2(V)α1(II) | Хрящи, стекловидное тело | Коллагенопатия II и XI типов, остеопороз |
| XII | COL12A1 | α1(XII)3 | Мягкие ткани | Повреждения сухожилий |
| XIII | COL13A1 | α1(XIII)3 | Поверхность клеток, эпителиальные клетки | |
| XIV | COL14A1 | α1(IV)3 | Мягкие ткани | |
| XV | COL15A1 | α1(XV)3 | Эндотелиальные клетки | Карцинома |
| XVI | COL16A1 | α1(XVI)3 | Повсеместно | |
| XVII | COL17A1 | α1(XVII)3 | Поверхность эпидермальных клеток | Буллезный эпидермиолиз, пузырчатка |
| XVIII | COL18A1 | α1(XVIII)3 | Эндотелиальные клетки | |
| XIX | COL19A1 | α1(XIX)3 | Повсеместно | Меланома, карцинома |
| XX | COL20A1 | α1(XX)3 | Выделен из куриного эмбриона | |
| XXI | COL21A1 | α1(XXI)3 | Кровеносные сосуды | |
| XXII | COL22A1 | α1(XXII)3 | Только в местах мышечно-сухожильных соединений | |
| XXIII | COL23A1 | α1(XXIII)3 | Опухолевые клетки | |
| XXIV | COL24A1 | α1(XXIV)3 | Формирующиеся кости | Остеохондроз |
| XXV | COL25A1 | α1(XXV)3 | Атеросклеротические бляшки | Болезнь Альцгеймера |
| XXVI | COL26A1=EMID2 | α1(XXVI)3 | Половые органы | |
| XXVII | COL27A1 | α1(XXVII)3 | Мягкие ткани | |
| XXVIII | COL28A1 | α1(XXVIII)3 | Нервная система |
Медицинские аспекты
Нарушения синтеза коллагена в организме лежат в основе таких наследственных заболеваний, как дерматоспораксис у животных, латиризм (характерна разболтанность суставов, привычные вывихи), синдром Элерса-Данлоса (до 14 типов проявлений), несовершенный остеогенез (болезнь «стеклянного человека», врождённый рахит, врождённая ломкость костей), болезнь Марфана.
Характерным проявлением этих заболеваний является повреждение связочного аппарата, хрящей, костной системы, наличие пороков сердечных клапанов.
Болезни, вызванные дефектами при биосинтезе коллагена, в том числе так называемые коллагенозы, возникают из-за множества причин. Это может быть из-за мутации в гене, кодирующем аминокислотную последовательность ферментов, продуцирующих коллаген, приводящей к изменению формы коллагеновой молекулы, или ошибки в посттранстляционной модификации коллагена. Также болезни могут быть вызваны недостатком или «неправильной работой» ферментов, вовлеченных в биосинтез коллагена — дефицит ферментов гидроксилирования (пролин-, лизингидроксилазы), гликозилтрансфераз, N-проколлагеновой и С-проколлагеновой пептидаз, лизилоксидаз с последующим нарушением поперечных сшивок, дефицит меди, витаминов В6, В13 (оротовая кислота), C. При приобретённых болезнях, таких как цинга, восстановление баланса ферментов до нормального может привести к полному излечению.
Практически любая генная мутация ведёт к утрате или изменению функций коллагена, что, в свою очередь, отражается на свойствах тканей и органов. Генные мутации в коллагеновом домене могут привести к изменению формы тройной спирали путём вставки/выпадения аминокислоты из полипептидной цепочки или замены Gly на другое основание. Мутации в неколлагеновых доменах могут привести к неправильной пространственной сборке α-цепей в надмолекулярные структуры (фибриллы или сети), что также ведёт к утрате функций. Мутантные α-цепи способны образовывать трёхспиральный комплекс с нормальными α-цепями. В большинстве случаев, такие комплексы нестабильны и быстро разрушаются, однако, такая молекула может и нормально выполнять свою роль, если не затронуты функционально важные области. Большинство болезней, вызванных мутациями в «коллагеновых» генах, являются доминантными.
Примечания
Ссылки
— простые белки — Биохимия
Структура простых белков представлена только полипептидной цепью (альбумин, инсулин). Однако необходимо понимать, что многие простые белки (например, альбумин) не существуют в «чистом» виде, они всегда связаны с какими-либо небелковыми веществами. Их относят к простым белкам только по той причине, что связи с небелковой группой слабые и при выделении in vitro они оказываются свободным от других молекул — простым белком.
Альбумины
Альбумины – это группа схожих белков плазмы крови с молекулярной массой 69 кДа, содержат много глутаминовой кислоты и поэтому имеют кислые свойства и высокий отрицательный заряд при физиологических рН. Легко адсорбируют полярные и неполярные молекулы, являются белком-транспортером в крови для многих веществ, в первую очередь для билирубина и длинноцепочечных жирных кислот.
В природе альбумины входят в состав не только плазмы крови (сывороточные альбумины), но и яичного белка (овальбумин), молока (лактальбумин), являются запасными белками семян высших растений.
Глобулины
Группа разнообразных белков плазмы крови с молекулярной массой до 100 кДа, слабокислые или нейтральные. Они слабо гидратированы, по сравнению с альбуминами меньше устойчивы в растворе и легче осаждаются, что используется в клинической диагностике в «осадочных» пробах (тимоловая, Вельтмана). Несмотря на то, что их обычно относят к простым, многие глобулины содержат углеводные или иные небелковые компоненты.
При электрофорезе глобулины сыворотки крови разделяются, как минимум, на 4 фракции – α1-глобулины, α2-глобулины, β-глобулины и γ-глобулины.
Картина электрофореграммы (вверху) белков сыворотки крови
и полученной на ее основе протеинограммы (внизу)
Так как глобулины включают в себя разнообразные белки, то их функции разнообразны:
Часть α-глобулинов обладает антипротеазной активностью, что защищает белки крови и межклеточного матрикса от преждевременного разрушения, например, α1-антитрипсин, α1-антихимотрипсин, α2-макроглобулин.
Некоторые глобулины способны к связыванию определенных веществ: трансферрин (переносит ионы железа), церулоплазмин (содержит ионы меди), гаптоглобин (переносчик гемоглобина), гемопексин (транспорт гема).
γ-Глобулины являются антителами и обеспечивают иммунную защиту организма.
Гистоны
Взаимодействие гистонов и ДНК |
Гистоны – внутриядерные белки массой около 24 кДа. Обладают выраженными основными свойствами, поэтому при физиологических значениях рН заряжены положительно и связываются с дезоксирибо-нуклеиновой кислотой (ДНК), образуя дезоксирибо-нуклеопротеины. Существуют 5 типов гистонов – очень богатый лизином (29%) гистон Н1, другие гистоны Н2а, h3b, НЗ, Н4 богаты лизином и аргинином (в сумме до 25%).
Радикалы аминокислот в составе гистонов могут быть метилированы, ацетилированы или фосфорилированы. Это изменяет суммарный заряд и другие свойства белков.
Можно выделить две функции гистонов:
1. Регуляция активности генома, а именно – они препятствуют транскрипции.
2. Структурная – стабилизируют пространственную структуру ДНК.
Гистоны в комплексе с ДНК образуют нуклеосомы – октаэдрические структуры, составленные из гистонов Н2а, h3b, НЗ, Н4. Гистон h2 связан с молекулой ДНК, не позволяя ей «соскользнуть» с гистонового октамера. ДНК обвивает нуклеосому 2,5 раза, после чего обвивает следующую нуклеосому. Благодаря такой укладке достигается уменьшение размеров ДНК в 7 раз.
Далее такие «бусы» нуклеосом могут складываться в суперспираль и в более сложную структуру нуклеопротеина.
Благодаря гистонам и формированию более сложных структур размеры ДНК, в конечном итоге, уменьшаются в тысячи раз: на самом деле длина ДНК достигает 6-9 см (10–1), а размеры хромосом – всего несколько микрометров (10–6).
Протамины
Это белки массой от 4 кДа до 12 кДа, имеются в ядрах сперматозоидов многих организмов, в сперме рыб (молóках) они составляют основную массу белка. Протамины являются заменителями гистонов и служат для организации хроматина в спермиях. По сравнению с гистонами протамины отличаются резко увеличенным содержанием аргинина (до 80%). Также, в отличие от гистонов, протамины обладают только структурной функцией, регулирующей функции у них нет, хроматин в сперматозоидах неактивен.
Коллаген
Коллаген – фибриллярный белок с уникальной структурой, составляет основу межклеточного вещества соединительной ткани сухожилий, кости, хряща, кожи, но имеется, конечно, и в других тканях.
Полипептидная цепь коллагена включает 1000 аминокислот и носит название α-цепь. Насчитывается около 30 вариантов α-цепи коллагена, но все они обладают одним общим признаком – в большей или меньшей степени включают повторяющийся триплет [Гли-Х-Y], где X и Y – любые, кроме глицина, аминокислоты. В положении X чаще находится пролин или, гораздо реже, 3-оксипролин, в положении Y встречается пролин и 4-оксипролин. Также в положении Y часто находится аланин, лизин и 5-оксилизин. На другие аминокислоты приходится около трети от всего количества аминокислот.
Жесткая циклическая структура пролина и оксипролина не позволяет образовать правозакрученную α-спираль, но образует т.н. «пролиновый излом». Благодаря такому излому формируется левозакрученная спираль, где на один виток приходится 3 аминокислотных остатка.
При синтезе коллагена первостепенное значение имеет гидроксилирование лизина и пролина, включенных в состав первичной цепи, осуществляемое при участии аскорбиновой кислоты. Также коллаген обычно содержит моносахаридные (галактоза) и дисахаридные (глюкоза-галактоза) молекулы, связанные с ОН-группами некоторых остатков оксилизина.
Этапы синтеза молекулы коллагена
Синтезированная молекула коллагена построена из 3 полипептидных цепей, сплетенных между собой в плотный жгут – тропоколлаген (длина 300 нм, диаметр 1,6 нм). Полипептидные цепи прочно связаны между собой через ε-аминогруппы остатков лизина. Тропоколлаген формирует крупные коллагеновые фибриллы диаметром 10-300 нм. Поперечная исчерченность фибриллы обусловлена смещением молекул тропоколлагена друг относительно друга на 1/4 их длины.
Фибриллы коллагена очень прочны, они прочнее стальной проволоки равного сечения. В коже фибриллы образуют нерегулярно сплетенную и очень густую сеть. Например, выделанная кожа представляет собой почти чистый коллаген.
Гидроксилирование пролина осуществляет железо-содержащий фермент пролилгидроксилаза для которого необходим витамин С (аскорбиновая кислота). Аскорбиновая кислота предохраняет от инактивации пролилгидроксилазу, поддерживая восстановленное состояние атома железа в ферменте. Коллаген, синтезированный в отсутствии аскорбиновой кислоты, оказывается недостаточно гидроксилированным и не может образовывать нормальные по структуре волокна, что приводит к поражению кожи и ломкости сосудов, и проявляется как цинга.
Гидроксилирование лизина осуществляет фермент лизилгидроксилаза. Она чувствительна к влиянию гомогентизиновой кислоты (метаболит тирозина), при накоплении которой (заболевания алкаптонурия) нарушается синтез коллагена, и развиваются артрозы.
Время полужизни коллагена исчисляется неделями и месяцами. Ключевую роль в его обмене играет коллагеназа, расщепляющая тропоколлаген на 1/4 расстояния с С-конца между глицином и лейцином.
По мере старения организма в тропоколлагене образуется все большее число поперечных связей, что делает фибриллы коллагена в соединительной ткани более жесткими и хрупкими. Это ведет к повышенной ломкости кости и снижению прозрачности роговицы глаза в старческом возрасте.
В результате распада коллагена образуется гидроксипролин. При поражении соединительной ткани (болезнь Пейджета, гиперпаратиреоидизм) экскреция гидроксипролина возрастает и имеет диагностическое значение.
Эластин
Строение десмозина |
По строению в общих чертах эластин схож с коллагеном. Находится в связках, эластичном слое сосудов. Структурной единицей является тропоэластин с молекулярной массой 72 кДа и длиной 800 аминокислотных остатков. В нем гораздо больше лизина, валина, аланина и меньше гидроксипролина. Отсутствие пролина обусловливает наличие спиральных эластичных участков.
Характерной особенностью эластина является наличие своеобразной структуры – десмозина, который своими 4-мя группами объединяет белковые цепи в системы, способные растягиваться во всех направлениях.
α-Аминогруппы и α-карбоксильные группы десмозина включаются в пептидные связи одной или нескольких белковых цепей.
Роль десмозина в соединении белков
Виды коллагена
Коллаген — полиморфный белок, в настоящее время известно 19 типов коллагена, которые отличаются друг от друга по первичной структуре пептидных цепей, функциям и локализации в организме. 95% всего коллагена в организме человека составляют коллагены I, II и III типов.
Типы | Гены | Ткани и органы |
I | COLIA1, COL1A2 | Кожа, сухожилия, кости, роговица, плацента, артерии, печень, дентин |
II | COL2A1 | Хрящи, межпозвоночные диски, стекловидное тело, роговица |
III | C0L3A1 | Артерии, матка, кожа плода, строма паренхиматозных органов |
IV | COL4A1-COL4A6 | Базальные мембраны |
V | COL5A1-COL5A3 | Минорный компонент тканей, содержащих коллаген I и II типов (кожа, роговица, кости, хрящи, межпозвоночные диски, плацента) |
VI | COL6A1-COL6A3 | Хрящи, кровеносные сосуды, связки, кожа, матка, лёгкие, почки |
VII | COL7A1 | Амнион, кожа, пищевод, роговица, хорион |
VIII | COL8A1-COL8A2 | Роговица, кровеносные сосуды, культуральная среда эндотелия |
IX | COL9A1-COL9A3 | Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
X | COL10A1 | Хрящи (гипертрофированные) |
XI | COLUA1-COL11A2 | Ткани, содержащие коллаген II типа (хрящи, межпозвоночные диски, стекловидное тело) |
XII | COL12A1 | Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XIII | C0L13A1 | Многие ткани |
XIV | COL14A1 | Ткани, содержащие коллаген I типа (кожа, кости, сухожилия и др.) |
XV | C0L15A1 | Многие ткани |
XVI | COL16A1 | Многие ткани |
XVII | COL17A1 | Гемидесмосомы кожи |
XVIII | COL18A1 | Многие ткани, например печень, почки |
XIX | COL19A1 | Клетки рабдомиосаркомы |
Гены коллагена называются по типам коллагена и записываются арабскими цифрами, например СОL1 — ген коллагена 1 типа, COL2 — ген коллагена II типа и т.д. К этому символу приписываются буква А (обозначает α-цепь) и арабская цифра (обозначает вид α-цепи). Например, COL1A1 и COL1A2 кодируют, соответственно, α1, и α2-цепи коллагена I типа.
Этапы синтеза и созревания коллагена
Синтез и созревание коллагена — сложный многоэтапный процесс, начинающийся в клетке, а завершающийся в межклеточном матриксе:
1. На полисомах ЭПР синтезируются полипептидные препро-α-цепиколлагена. Они содержит начиная сN-конца: 1). гидрофобный «сигнальный» пептид, содержащий около 100 АК; 2). N-концевой пропептид, содержащий около 100 АК, в том числе цистеин; 3). α-цепь коллагена 4). С-концевой пропептид, содержащий около 250 АК, в том числе цистеин. КонцевыеС- и N-пропептидыформируют глобулярные домены и необходимы для правильного формирования тройной спирали.«Сигнальный» пептид, обеспечивает поступление синтезируемой на рибосоме препро-α-цепи в полость ЭПР.
2. В полости ЭПР при отщеплении сигнального пептида препро-α-цепиколлагена превращаются впро-α-цепи.
3. Поступающие в полости ЭПР про-α-цепи коллагенаподвергаются модификации.
а). Цистеины N-пропептидов образуют внутрицепочечные дисульфидные мостики, формируя наN-конце глобулярную структуру;
б). Пролины и лизины в Y-положении (гли-х-у) про-α-цепей гидроксилируются пролил-4-гидроксилазойилизил-5-гидроксилазойв 4-гидроксипролины (Hyp) и 5-гидроксилизины (Hyl). Некоторые пролины в Х-положениях гидроксилируются в 3-гидроксипролины пролил-3-гидроксилазой. Оксигеназы (гидроксилазы), содержат Fe2+, находятся на мембране ЭПР. Для реакции необходимы а-КГ, О2и витамин С:
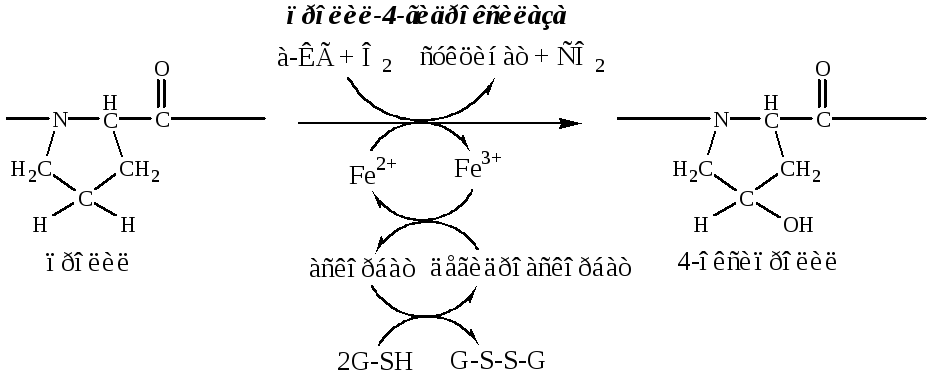
Гидроксилирование пролина необходимо для стабилизации тройной спирали коллагена, ОН-группы гидроксипролина участвуют в образовании водородных связей.
Гидроксилирование лизина необходимо для последующего образования ковалентных связей между молекулами коллагена при сборке коллагеновых фибрилл.
в). Гидроксилизин про-α-цепей при участии гликозилтрансфераз гликозилируется галактозой или галактозилглюкозой. В молекуле коллагена сухожилий (тип I) количество углеводов равно 6, а в моллекуле коллагена капсулы хрусталика (тип TV) — 110. Роль этих углеводных групп неясна.
4). В просвете ЭПР после отделения от рибосом про-α-цепей, 3 из них с помощью С-концевых пропептидов соединяются между собой дисульфидными мостиками (цистеины С-пропептидов образуют внутри- и межцепочечные дисульфидные мостики) и скручиваются с образованием тройной спирали проколлагена. Тройная спираль проколлагена стабилизируется водородными связями. После этого гидроксилирование и гликозилирование про-α-цепей прекращается.
5). Из ЭПР молекулы проколлагена перемещаются в аппарат Гольджи, включаются в секреторные пузырьки и секретируются в межклеточное пространство.
6). В межклеточном матриксе от некоторых проколлагенов(I, II, III, V, XI типов)проколлагенпептидазыотщепляют концевые С- иN-пропептиды, в результате чего образуетсятропоколлагены. У проколлагенов IV, VIII, X типов концевые пропептиды не отщепляются.
Синтезированные молекулы проколлагенов и тропоколлагенов способны образовывать различные структуры. Например, тропоколлагены образуют фибриллы, а проколлагены образуют сети.
Структура | Тип коллагена |
Фибриллы | I, II, III – основные. V, XI — минорные |
Ассоциированные с фибриллами | IX, XII, XIV, XVI, XIX |
Сети | IV, VIII, X |
Микрофибриллы | VI |
«Заякоренные» фибриллы | VII |
Трансмембранные домены | XIII, XVII |
Другие | XV, XVIII |